Anatomy and Structural Biology, PhD and MSc Programs.

กตัญญู.. ต่อครูผู้ยิ่งใหญ่

-

นักศึกษาแพทย์โครงการผลิตแพทย์เพื่อชาวชนบท ชั้นปีที่ 2 มหาวิทยาลัยมหิดล จัดงานอำลาร่างอาจารย์ใหญ่ประจำปีการศึกษา 2566
-

ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล ร่วมกับนักศึกษาวิทยาศาสตร์สุขภาพ จัดงานทำบุญอุทิศส่วนกุศลแด่อาจารย์ใหญ่ วิทยาเขตศาลายา
-

เพจนิรันทร์ เพจห่มบุญ และเมืองสมุยกรุ๊ป ร่วมสนับสนุนการดูแลร่างอาจารย์ใหญ่ของภาควิชากายวิภาคศาสตร์ด้วยผ้าขาวห่อร่าง และถุงซิปบรรจุร่างอาจารย์ใหญ่
-

ขอเชิญร่วมพิธีทำบุญอุทิศส่วนกุศลแด่ผู้อุทิศร่างกายเพื่อการศึกษา ประจำปีการศึกษา ๒๕๖๖
SDGs & Research output (ผลงานวิจัย)
Anatomical study and meta-analysis of the episternal ossicles
Highlight การศึกษานี้ประกอบด้วยการศึกษาจากภาพเอกซเรย์คอมพิวเตอร์ (computed tomography) และการวิเคราะห์อภิมานจากฐานข้อมูลอิเล็กทรอนิก 3 แหล่งได้แก่ Google Scholar, PubMed, and Journal Storage ผลงานวิจัยพบว่า จากคนไข้จำนวน 7,997 รายจากผลงานตีพิมพ์กว่า 16 ฉบับ episternal ossicles มีความชุกอยู่ที่ 2.1% โดยการศึกษาด้วย X-ray มีความชุกสูงสุดอยู่ที่ 7% และประชากรเอเชียมีความชุกของ episternal ossicles […]
-
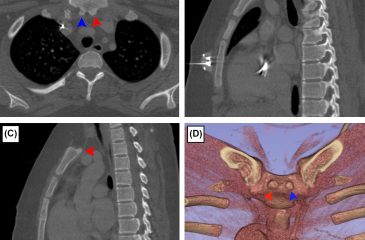
Radiological Study of Atlas Arch Defects with Meta-Analysis and a Proposed New Classification
-

Normative Reference Values of the Tibial Nerve in Healthy Individuals Using Ultrasonography: A Systematic Review and Meta-Analysis
-
Basilar tubercles and eminences of the clivus: Novel anatomical entities
-
Extracts of tropical green seaweed Caulerpa lentillifera reduce hepatic lipid accumulation by modulating lipid metabolism molecules in HepG2 cells
-
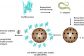
Biosynthesis of Cry5B-Loaded Sulfur Nanoparticles using Arthrobotrys oligospora Filtrate: Effects on Nematicidal Activity, Thermal Stability, and Pathogenicity against Caenorhabditis elegans
-
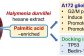
Apoptotic and Autophagic Cell Death Effects of the Hexane Extract of Tropical Marine Algae Halymenia durvillei against Human Glioblastoma Cells: In vitro and in silico Studies
-
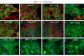
Unveiling the Antiviral Properties of Panduratin A through SARS-CoV-2 Infection Modeling in Cardiomyocytes
-

Octanoyl esterification of low molecular weight sulfated galactan enhances the cellular uptake and collagen expression in fibroblast cells
-

The Anatomical Relationship Between the Cervical Nerve Roots, Intervertebral Discs and Bony Cervical Landmark for Posterior Endoscopic Cervical Foraminotomy and Discectomy: A Cadaveric Study
University Life

Salinthip Thongdechsri’s Ph.D. Proposal Examination on Thursday 25th April 2024, 09:00-12:00, @AN1-202. All are welcome.
Effects of Holothuria leucospilota Extracts and Bioactive Compounds on Amyloid Beta Aggregation and Its Toxicity in Transgenic Caenorhabditis elegans
-

Warachin Gangnonngiw’s Ph.D. Thesis Defense on Thursday 18th April 2024, 09:00-12:00, @AN1-202. All are welcome.
-

Chidchanok Chawiwithaya’s Ph.D. Proposal Examination on Thursday 11th April 2024, 09:00-12:00, @AN1-205. All are welcome.
-

Chanathip Kamonnat’s M.Sc. Proposal Examination on Thursday 7th March 2024, 09:00-12:00, @AN1-202. All are welcome.
-

Sukanya Saedan’s Ph.D. Qualifying Examination on Tuesday 5th March 2024, 09:00-12:00, @AN1-202. All are welcome.
Scholarship

เรียนเชิญร่วมบริจาคสมทบทุนเพื่อจัดซื้อครุภัณฑ์ทางการศึกษาอาจารย์ใหญ่
เนื่องด้วย ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล มีความประสงค์ที่จะจัดซื้อครุภัณฑ์สำหรับการเรียนปฏิบัติการร่างอาจารย์ใหญ่และการฝึกผ่าตัดเสมือนจริงในอาจารย์ใหญ่เพื่อเพิ่มพูนความรู้ความสามารถและประสบการณ์ให้แก่นักศึกษาแพทย์และแพทย์เฉพาะทาง ภาควิชากายวิภาคศาสตร์จึงใคร่ขอเรียนเชิญบริจาคสมทบทุนเพื่อจัดซื้อครุภัณฑ์ทางการศึกษาอาจารย์ใหญ่ ดังรายการต่อไปนี้ 1. โคมไฟผ่าตัด LED แบบเคลื่อนที่ 40,000 บาท จำนวน 5 ทุน 2. เครื่องฉีดน้ำยารักษาสภาพร่างอาจารย์ใหญ่แบบนิ่มเสมือนจริง 500,000 บาท จำนวน 1 ทุน * หมายเหตุ สามารถบริจาคได้มากกว่า 1 ทุน หรือ หลายท่านบริจาคร่วมกันเป็น 1 ทุน นิติบุคคล/บุคคลธรรมดา สามารถบริจาค […]
-

(ขอปิดรับการบริจาคทุนฯ เนื่องจากมีผู้แจ้งความประสงค์บริจาคเงินครบตามจำนวนทุนการศึกษาแล้ว) เรียนเชิญบริจาคทุนการศึกษาเพื่อจัดสรรให้นักศึกษาแพทย์ที่ขาดแคลนทุนทรัพย์ ปี 2567
-

ประกาศรายชื่อนักศึกษาแพทย์ผู้ที่มีสิทธิ์เข้ารับการสัมภาษณ์เพื่อรับทุนการศึกษา จากผู้บริจาคทุน จัดสรรโดยภาควิชากายวิภาคศาสตร์ ปีการศึกษา 2567
Thiel cadaver

ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Thiel cadaver) ครั้งที่ 2/2567
เมื่อวันที่ 21 มีนาคม 2567 ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่นายสำรวย อินทร์ยงค์ โดยมีญาติอาจารย์ใหญ่และเจ้าหน้าที่ของภาควิชาฯเข้าร่วมพิธี ณ วัดสุคันธาราม เขตดุสิต กรุงเทพฯ เวลา 10.00 น. การอุทิศร่างกายของอาจารย์ใหญ่ในครั้งนี้ภาควิชาฯได้ศึกษาร่างอาจารย์ใหญ่เพื่อการศึกษาทางการแพทย์และเพื่องานวิจัยทางด้านกายวิภาคศาสตร์ในการหาองค์ความรู้ใหม่ เพื่อพัฒนาการเรียนการสอนกายวิภาคศาสตร์สำหรับนักศึกษาแพทย์ และวิชาการการแพทย์ โดยในครั้งนี้ ภาควิชากายวิภาคศาสตร์ฯ ได้ดำเนินการศึกษาและวิจัย ร่วมกับภาควิชากายวิภาคศาสตร์ คณะแพทยศาสตร์ มหาวิทยาลัยขอนแก่น และ University of Birmingham […]

Soft cadaver คือ ร่างของผู้บริจาคร่างกายเพื่อการศึกษาแบบนิ่ม ซึ่งเสมือนกับร่างของผู้ป่วยจริงมากที่สุด

ภาควิชากายวิภาคศาสตร์ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Soft cadaver) ครั้งที่ 12/2567
เมื่อวันที่ 18 มีนาคม 2567 ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่นายเกษมสันติ์ กีรติเลิศบุญ โดยมีญาติอาจารย์ใหญ่และเจ้าหน้าที่ของภาควิชาฯเข้าร่วมพิธี ณ วัดสุคันธาราม เขตดุสิต กรุงเทพฯ เวลา 10.00 น. การอุทิศร่างกายของอาจารย์ใหญ่เพื่อการศึกษาทางการแพทย์โดยการใช้ร่างเพื่อฝึกผ่าตัดผ่านกล้อง ในครั้งนี้เป็นของสมาคมศัลยแพทย์ทรวงอกแห่งประเทศไทย เพื่อให้เกิดความชำนาญในการนำความรู้ไปใช้รักษาผู้ป่วยต่อไป ทั้งนี้ภาควิชาฯ จะนำเถ้ากระดูก เล็บและเส้นผมของอาจารย์ใหญ่เข้าร่วมพิธีพระราชทานเพลิงศพฯ ต่อไป ขอบุญกุศลที่ท่านอาจารย์ใหญ่ได้อุทิศไว้ เป็นผลบุญนำดวงวิญญาณของท่านอาจารย์ใหญ่สู่สุขคติในสัมปรายภพ ที่ปรึกษา การเขียนข่าวเพื่อประชาสัมพันธ์: รศ.ดร.สมลักษณ์ อสุวพงษ์พัฒนาเขียนข่าว: […]
-

ภาควิชากายวิภาคศาสตร์ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Soft cadaver) ครั้งที่ 11/2567
-

ภาควิชากายวิภาคศาสตร์ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Soft cadaver) ครั้งที่ 10/2567
-

ภาควิชากายวิภาคศาสตร์ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Soft cadaver) ครั้งที่ 9/2567
-

ภาควิชากายวิภาคศาสตร์ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Soft cadaver) ครั้งที่ 8/2567
-

ภาควิชากายวิภาคศาสตร์ได้ดำเนินการฌาปนกิจร่างอาจารย์ใหญ่ (Soft cadaver) ครั้งที่ 7/2567
Activities (กิจกรรม)
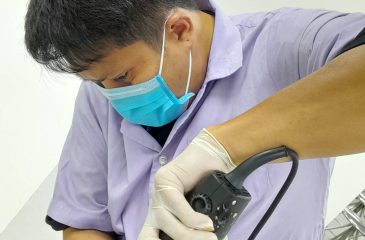
เจ้าหน้าที่รักษาร่างอาจารย์ใหญ่ ภาควิชากายวิภาคศาสตร์ ฝึกทักษะการเปิดกะโหลกศรีษะด้วยเลื่อยไฟฟ้า
ด้วยภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล รับผิดชอบการเรียนการสอนรายวิชามหกายวิภาคศาสตร์ให้กับนักศึกษาแพทย์ ในโครงการผลิตแพทย์เพื่อชาวชนบท มหาวิทยาลัยมหิดล ที่ยังคงใช้การเปิดกระโหลกศรีษะอาจารย์ใหญ่ด้วยเลื่อยมือ และได้เห็นว่าการใช้เลื่อยไฟฟ้าจะทำให้ประหยัดเวลาในการปฏิบัติมากกว่า ซึ่งการใช้เลื่อยไฟฟ้าเปิดกระโหลกศรีษะ ต้องมีทักษะในการลงน้ำหนักเพื่อไม่ให้ตัดโดนเยื่อหุ้มสมอง ในการนี้ภาควิชาฯ จึงได้ส่ง 4 เจ้าหน้าที่ ได้แก่ นายศิวา สุขประเสริฐ นายภานุวัฒน์ ไพรวัลย์ นายศุภกิจ รักใคร่ และนายนันทาภิวัฒน์ โพธิพิพัฒนกุลโดยได้รับความอนุเคราะห์จาก ผู้ช่วยศาสตราจารย์ ดร.เดชา บูรณจิตร์ภิรมย์ หัวหน้าสาขากายวิภาคศาสตร์ คณะแพทยศาสตร์ มหาวิทยาลัยธรรมศาสตร์ ให้ความอนุเคราะห์การฝึกทักษะจากผู้เชี่ยวชาญคุณวัชรเทพ […]
-

ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล ให้การต้อนรับอาจารย์ และบุคลากร จาก Universitas Negeri Yogyakarta ประเทศอินโดนีเซีย เข้าศึกษาดูงาน การจัดการเรียนการสอนกายวิภาคศาสตร์ ณ ห้องปฏิบัติการกายวิภาคศาสตร์ วิทยาเขตศาลายา
-

SPECIAL SEMINAR IN ANATOMY: Potential use of native and engineered cathelicidin peptides as vaginal contraceptives and anti-infectives against sexually transmitted infection (STI) microbes by Emeritus Professor Nongnuj Tanphaichitr – All are welcome
-

กิจกรรมส่งท้ายปีเก่าต้อนรับปีใหม่ 2567
-

ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล เป็นฐานการจัด Workshop Fundamental Blepharoplasty และ การฝึกหัตถการการผ่าตัดตา 2 ชั้น
Easy IT

คู่มือ การใช้คอมพิวเตอร์ (Main PC) ประจำห้องเรียน SC2-152 ประกอบการบันทึกการสอนผ่าน Webex
กรณีที่ 1: ใช้ Main PC ในการสอน สำคัญมาก: ไฟสีฟ้า ต้องอยู่ที่ปุ่ม Main PC เท่านั้น หมายเหตุ โดยปกติแล้ว เจ้าหน้าที่ฯ จะเปิดเครื่องคอมพิวเตอร์ (Main PC) ไว้ให้ก่อนมีการเรียนการสอนถ้าหากพบว่า เครื่อง Main PC ยังไม่เปิด ให้ ดำเนินการเปิดเครื่องได้ทันที หรือติดต่อเจ้าหน้าที่โสตทัศนศึกษา งานการศึกษา ศาลายาเบอร์ 02 441 […]
-

Anatomy: ON-LINE IT services
-

บันทึกหน้าจอด้วย Power Point 2019
-
การ join Webex Meeting ด้วย web browser
-
การขอสิทธิ์เข้าใช้งาน Webex Meeting (สำหรับอาจารย์และเจ้าหน้าที่)
Staff Awards & Promotions

ขอแสดงความยินดีกับ ผู้ช่วยศาสตราจารย์ ดร.นพพร จงกมลวิวัฒน์ อาจารย์ประจำภาควิชากายวิภาคศาสตร์
เนื่องในโอกาสที่ได้รับการอนุมัติแต่งตั้งจากสภามหาวิทยาลัยมหิดลให้ดำรงตำแหน่ง รองศาสตราจารย์ สาขาวิชาประสาทวิทยาศาสตร์ ตั้งแต่ วันที่ 21 มิถุนายน 2565 ตามมติที่ประชุมสภามหาวิทยาลัย ในการประชุมครั้งที่ 600 วันที่ 17 มกราคม 2567 เครดิตภาพ: คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล เว็บมาสเตอร์: ว่าที่ ร.อ. นเรศ จันทรังสิกุล
-

Congratulations! to Head of Department, Assoc. Prof. Dr. Wattana Weerachatyanukul
-

ขอแสดงความยินดีกับ ผู้ช่วยศาสตราจารย์ ดร.วรวิทย์ ศุภมั่งมี อาจารย์ประจำภาควิชากายวิภาคศาสตร์
Congratulations!!

Proudly congratulate to new Dr. Boottoh Nambunruang after successfully defending his dissertation
Boottoh Nambunruang, a doctoral candidate in our department, successfully defended his dissertation on Thursday 28th December 2023. The title of his work is, “Effect of sulfated galactans on chondrocyte functions […]
-

Proudly congratulate to new Dr. Kanitin Rumpansuwon after successfully defending his dissertation
-

Proudly congratulate to new Dr. Jinchutha Duangdara after successfully defending her dissertation
SDGs & Activity/Project (กิจกรรม/โครงการ)

นักศึกษาแพทย์โครงการผลิตแพทย์เพื่อชาวชนบท ชั้นปีที่ 2 มหาวิทยาลัยมหิดล จัดงานอำลาร่างอาจารย์ใหญ่ประจำปีการศึกษา 2566
เมื่อวันเสาร์ที่ 9 มีนาคม พ.ศ. 2567 เวลา 8.30-9.30 น. ณ ห้องปฏิบัติการกายวิภาคศาสตร์ AN1-301 ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล นักศึกษาแพทย์โครงการผลิตแพทย์เพื่อชาวชนบท ศูนย์แพทยศาสตร์ศึกษาโรงพยาบาลมหาราช จ.นครราชสีมา ศูนย์แพทยศาสตร์ศึกษาโรงพยาบาลสวรรค์ประชารักษ์ จังหวัดนครสวรรค์ ศูนย์แพทย์ศาสตร์ศึกษาโรงพยาบาลมหาราช จ.นครศรีธรรมราช พร้อมด้วยคณาจารย์และเจ้าหน้าที่ ภาควิชากายวิภาคศาสตร์ ร่วมกันจัดงานอำลาร่างอาจารย์ใหญ่ประจำปีการศึกษา 2566 จำนวน 29 ท่าน โดยพิธีเริ่มด้วยการกล่าวบทขอขมาและบูชาคุณของอาจารย์ใหญ่ ตามด้วยการกล่าวความรู้สึกของตัวแทนนักศึกษาแพทย์ […]
-

การใช้ร่างอาจารย์ใหญ่สูตร Thiel ร่วมกับคณะแพทยศาสตร์โรงพยาบาลรามาธิบดีและคณะวิศวกรรมศาสตร์ มหาวิทยาลัยมหิดล
-

นักศึกษาแพทย์ ชั้นปีที่ 2 โครงการผลิตแพทย์เพื่อชาวชนบท จัดพิธีอำลาร่างอาจารย์ใหญ่ ประจำปีการศึกษา 2565
-

ภาควิชากายวิภาคศาสตร์ จัด Post-Conference: Cadaveric Course ให้กับสมาคมแพทย์ความงาม (ATAP)
-

มหิดลวิชาการ คณะวิทยาศาสตร์ ปี 2564
รับสมัครงาน 2 อัตรา

รับสมัครงาน ภาควิชากายวิภาคศาสตร์ คณะวิทยาศาสตร์ มหาวิทยาลัยมหิดล จำนวน 2 อัตรา
Link: แบบสำรวจความต้องการเข้าสมัครงาน หมายเหตุ ***Link แบบฟอร์มนี้เป็นเพียงแบบสำรวจความต้องการเข้าสมัครงาน และให้ข้อมูลเบื้องต้นแก่ภาควิชาฯ ยังไม่ใช่แบบฟอร์มการรับสมัครงาน ผู้ตอบแบบสอบถามจำเป็นต้องติดตามประกาศการรับสมัครงานผ่านระบบของมหาวิทยาลัยและสมัครผ่านระบบของมหาวิทยาลัย*** ติดตามประกาศรับสมัครงานได้ที่ https://muhr.mahidol.ac.th/E-Recruitment/job.php ตำแหน่งที่ 1: นักวิทยาศาสตร์ จำนวน 1 อัตรา คุณสมบัติทั่วไป 1.สำเร็จการศึกษาระดับปริญญาตรีหรือสูงกว่าจากมหาวิทยาลัยที่กรรมการบริหารมหาวิทยาลัยมหิดลให้การรับรองในสาขาทางวิทยาศาสตร์ หากเป็นสาขาที่มีการเรียนวิชากายวิภาคศาสตร์จะได้รับการพิจารณาเป็นพิเศษ 2.มีความรู้ความสามารถในการใช้คอมพิวเตอร์พื้นฐานในโปรแกรม Microsoft office และ Web based application ทางการเรียนการสอน เช่น Google Classroom 3.มีทักษะด้านการวิจัย […]